
Conformation of Ultra-Long Chain Fatty Acid in Lipid Bilayer
PI of Joint-use project: K. Kawaguch
Host lab: Noguchi Group
Host lab: Noguchi Group
Phospholipids are the most abundant lipids in biological membranes and have a polar head group and two hydrocarbon tails (fatty acids). Phospholipids, more than 1000 molecular species, are biosynthesized by a combination of a head group and two hydrocarbon tails. Each tail typically contains between 14 and 22 carbon atoms. Fatty acids containing more than 22 carbons are called very-long-chain fatty acids, such as docosahexaenoic acid (DHA, C22:6). Moreover, much longer chains containing more than 32 carbons were found at the sn-1 position of phosphatidylcholine in photoreceptors, fibroblasts, and keratinocytes. These fatty acids are called ultra-long-chain fatty acids (ULCFAs). It is considered that ULCFAs are stored as a precursor of bioactive lipid mediators and derivatives of C32:6 and C34:6 are neuroprotective in retina. However, the physicochemical properties of ULCFAs and the biological roles of ULCFAs-containing phospholipids are still unclear. In order to clarify the physicochemical properties of ULCFAs, we have simulated a single ULCFA in a lipid bilayer by all-atom molecular dynamics using the ISSP supercomputer [1,2].
We found that ULCFAs have large conformational fluctuations. Figure 1 shows the conformations of dotriacontahexaenoic acid containing phosphatidylcholine (dTSPC, C32:6-C18:0) in membrane of distearoyl phosphatidylcholine (DSPC, C18:0-C18:0) [1]. The ultra-long chain has a turned conformation in the upper leaflet, a L-shaped conformation between the two leaflets, and an elongated conformation deeply inserting in the lower (opposite) leaflet. The L-shaped conformation is taken most frequently. There are no free-energy barriers among these conformations and the transit time between turned and elongated conformations is ~ 10 ns. Moreover, we revealed that ULCFAs can sense the density differences between the two leaflets and respond to these changes. As the number ratio of lipids in the opposite leaflet increases, the ratio of the elongated shape linearly decreases. This change in ULCFA conformations reduces the lipid-density difference between the two leaflets. We have examined three types of phospholipids (DSPC, SDPC, C18:0-C22:6, and SOPC, C18:0-C18:1) for host membranes and three types of ULCFAs (dTSPC, HSPC, C26:4-C18:0, and LSPC, C32:0-C18:0) [1,2]. HSPC is constructed by truncating the long hydrocarbon chain of dTSPC at C26, and LSPC is constructed by the saturation of the long hydrocarbon chain of dTSPC. In all cases, ULCFAs have this sensing capability. Therefore, we concluded that it is a unique property of ULCFAs. This response may be essential for the functions of ULCFAs in living cells.
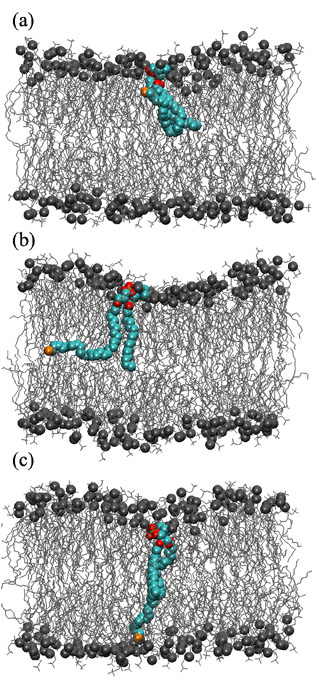
Fig. 1. Snapshots of a Ultra-Long Chain Fatty Acid, dTSPC, in bilayer membrane. (a) Turned conformation. (b) L-shaped conformation. (c) Elongated conformation. dTSPC are represented by colored spheres. Host lipid (DSPC) molecules are shown in gray (spheres represent the phosphate atoms). Water molecules are not shown for clarity.
Moreover, we added cholesterols to membranes [2]. The flip—flop of cholesterols is ~1 μs and is much faster than those of phospholipids (hours or days). The flip—flop of cholesterols can also reduce the lipid-density difference. After the equilibration of cholesterols, the probability distribution of ULCFA conformations is back to that for no-lipid-density-difference membranes. Thus, we consider that ULCFAs initially respond to the density difference and, later, cholesterol removes the difference in living cells.
References
- [1] K. Kawaguchi, K. M. Nakagawa, S. Nakagawa, H. Shindou, H. Nagao, and H. Noguchi, J. Chem. Phys. 153, 165101 (2020).
- [2] K. Kawaguchi, H. Nagao, H. Shindou, and H. Noguchi, J. Phys. Chem. B 126, 9316 (2022).