
Morphology of High-Genus Fluid Vesicles
Noguchi Group
The nucleus of a eukaryotic cell is surrounded by a nuclear envelope. The nuclear envelope consists of two bilayer membranes connected by many lipidic pores, which are supported by a protein complex called nuclear pore complex (NPC). Nuclear pores have an approximately uniform distribution in the nuclear envelope. Hence, the nuclear envelope is a spherical stomatocyte with a high genus. In order to clarify the formation mechanism of the nuclear envelope, we simulated the morphology of high-genus vesicles by dynamically triangulated membrane methods.
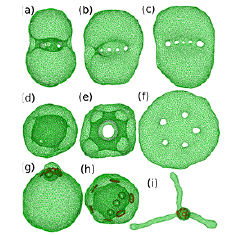
Fig. 1. Snapshots of high-genus vesicles (a–f) with no pore-size constraint and (g–h) with pore-size constraint. (a–g) g = 5. (h,i) g = 8.
First, we investigated the vesicle morphology of genus 0≦g≦8 in the absence of NPCs (see Fig. 1(a)–(f)) [1]. For g≧3, bending-energy minimization without volume or other constraints produces a circular-cage stomatocyte, where the pores are aligned in a circular line on an oblate bud (see Fig. 1(a)). As osmotic pressure is imposed to reduce the vesicle (perinuclear) volume, the vesicle transforms to the spherical stomatocyte (nuclear envelope shape) (see Fig. 1(d)). In the lipid vesicles, the area difference ΔA of two monolayers of the bilayer is different from the preferred value ΔA0 determined by the lipid number of both monolayers. This effect is taken into account by the area-difference elasticity (ADE) energy: Kade(ΔA−ΔA0)2/2. With increasing ΔA0, the vesicle transforms from the circular-cage stomatocyte into discocyte continuously via pore opening (see Fig. 1(b)) at the large volume as seen in genus-0 vesicles. Surprisingly, however, at the small volume, the vesicle exhibits a discrete transition from polyhedron to discocyte (see Figs. 1(e) and (f)). Thus, the pore arrangements change the character of shape transitions.
Next, we modeled the pore size constraint by the NPC as a ring which the membrane cannot penetrate [2]. When the pore is restricted as a small size, the aligned pores move to the end of the vesicle under the bending-energy minimization as shown in Fig. 1(g).
Interestingly, the pore-constraint itself rather presents the formation of the spherical stomatocyte. We found that the spherical stomatocyte is formed by a small perinuclear volume, osmotic pressure within nucleoplasm, and/or repulsion between the pores (see Fig. 1(h)). We consider the osmotic pressure by nucleosomes and nuclear proteins in nucleoplasm is the main source to stabilize the nuclear shape. When the ADE energy is accounted, the endoplasmic-reticulum-like tubules can grow from the spherical stomatocyte (see Fig. 1(i)).
References
- [1] H. Noguchi, EPL 112, 58004 (2015).
- [2] H. Noguchi, Biophys. J. 111, 824 (2016).